
As key regulators of innate immune responses and tissue homeostasis, macrophages possess prominent functional plasticity. Among various classification patterns, the polarization into M1 and M2 subtypes is the most well-documented classification system. The dynamic balance between these two polarized states serves as a core biological feature of macrophages, which profoundly affects disease progression, therapeutic efficacy and tissue regeneration processes, thus becoming a hot research topic in modern biomedicine. Clarifying the distinct characteristics of M1 and M2 macrophages contributes to revealing the pathogenesis of immune-related disorders, developing targeted therapeutic strategies and advancing overall immunological research progress.
This paper systematically elaborates the phenotypic traits, molecular features and biological functions of M1 and M2 macrophages, as well as their pathophysiological roles and updated research technologies.
Macrophages are not functionally fixed cell populations; instead, they can flexibly adjust their biological activities in response to diverse microenvironmental signals, and this functional transformation process is defined as cell polarization. Derived from peripheral blood monocytes or local tissue resident progenitor cells, macrophages are mainly divided into two typical functional subtypes: pro-inflammatory M1 macrophages and tissue-repair related M2 macrophages.
Established in the early 2000s, the M1/M2 classification model was inspired by the classification standard of Th1 and Th2 helper T cells. Classically activated M1 macrophages are mainly induced by IFN-γ, lipopolysaccharide (LPS) and other stimuli, acting as vital defensive barriers against pathogenic microorganisms and tumor lesions. These cells can secrete large quantities of pro-inflammatory mediators including TNF-α, IL-1β and IL-6, and generate reactive oxygen species (ROS) as well as reactive nitrogen species (RNS), further facilitating the activation of Th1-type immune responses to eliminate invasive pathogens and damaged somatic cells efficiently.
In contrast, alternative activated M2 macrophages are primarily induced by IL-4 and IL-13. They mainly take charge of inflammatory response subsidence, cellular debris clearance, tissue remodeling, wound healing, angiogenesis and immune regulation. Typical surface markers such as CD206 and ARG-1 are highly expressed on M2 macrophages, which can release anti-inflammatory cytokines like IL-10 and TGF-β. Furthermore, M2 macrophages can be further subdivided into four subgroups including M2a, M2b, M2c and M2d with differentiated functions.
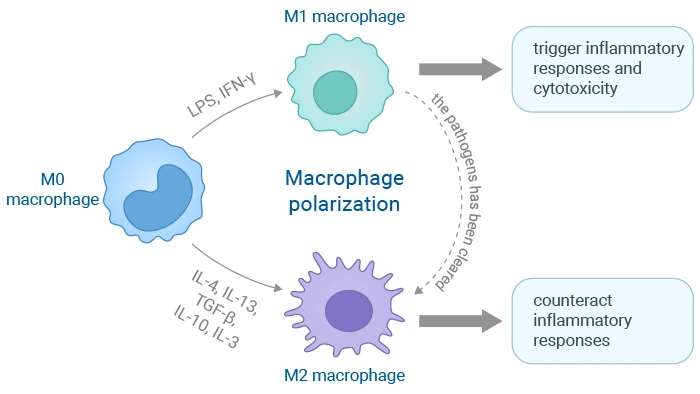
The functional differentiation of macrophages into classical M1 phenotype or alternative M2 phenotype is a key biological event involved in immune regulation, inflammatory reaction and tissue repair. Such functional switching behavior is precisely controlled by complicated intracellular signaling networks which can accurately respond to different microenvironmental cues.
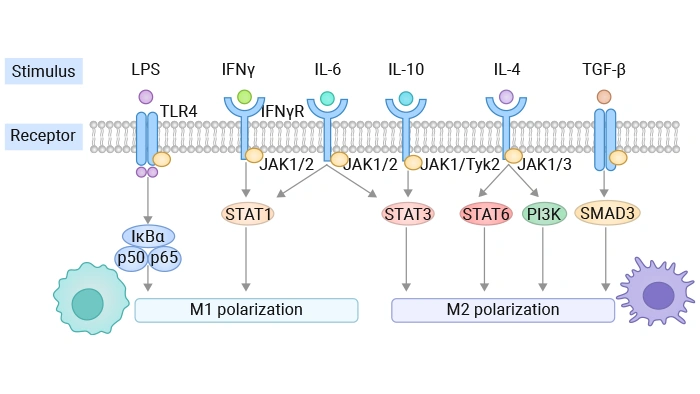
The NF-κB pathway acts as the dominant regulatory pathway controlling the transcription of M1-related pro-inflammatory genes. In most cases, extracellular stimuli such as LPS bind to pattern recognition receptors represented by TLR4 to activate this signaling cascade. Afterwards, inhibitory proteins including IκBα undergo phosphorylation and subsequent degradation, enabling nuclear translocation of NF-κB complexes represented by p65. After entering the nucleus, NF-κB can initiate the transcription of typical M1 characteristic genes such as iNOS, TNF-α, IL-6 and IL-1β.
The formation of M1 polarization state is usually co-regulated by JAK-STAT1 pathway together with NF-κB pathway. After IFN-γ binds to its corresponding membrane receptor, it can activate downstream JAK1 and JAK2 proteins, which further induce the phosphorylation modification of STAT1. Phosphorylated STAT1 forms homologous dimers and transfers into cell nucleus, binding to GAS sequence segments to start the expression of a series of M1 characteristic genes including IRF8, CIITA and GBP.
The combination between Notch ligands (Jagged1, Jagged2, DLL1, DLL4) on adjacent cells and macrophage surface Notch receptors (Notch1-4) can activate canonical Notch signaling. After signal activation, intracellular NICD segments are cleaved and released, which bind to nuclear transcription factor RBP-J to regulate downstream gene transcription. This pathway can amplify TLR-mediated signal transmission, directly upregulate Nos2 gene expression, and synergize with NF-κB to elevate the expression level of IL-6 and IL-12, ultimately driving macrophages to polarize towards M1 phenotype. Meanwhile, NICD can interact with HIF-1α to regulate glycolytic metabolism level during M1 cell activation.
When IL-4 or IL-13 bind to their respective membrane receptors, they can sequentially activate downstream JAK family kinases, which trigger STAT6 phosphorylation and dimerization process. Nucleus-located phosphorylated STAT6 can activate the transcription of classic M2a marker genes such as Arg1, Fizz1, Ym1 and Mrc1 (CD206). In addition, STAT6 can cooperate with transcription factor PPAR-γ and induce IRF4 expression, which further strengthens M2a characteristic gene expression and simultaneously suppresses the transcription of M1-related genes.
The binding reaction between IL-10 and its specific receptor can activate JAK1 and TYK2, leading to STAT3 phosphorylation and dimerization. Activated STAT3 can initiate the transcription of multiple M2c macrophage related genes including IL-10, TGF-β, Arg1, HIF2α and SOCS3, and effectively restrain the synthesis and release of various pro-inflammatory cytokines.
After TGF-β binds to cell surface receptors, it promotes the phosphorylation of receptor-regulated SMAD2 and SMAD3. These phosphorylated proteins form compound complexes with SMAD4 and enter the nucleus to regulate target gene expression. This pathway mainly regulates the synthesis of extracellular matrix components, fibronectin and PAI-1, and participates in mediating the fibrogenic effect and tissue remodeling function of M2 macrophages, which is of great significance in wound healing and tissue fibrosis research.
Activated mainly by IL-4 through IRS-2 molecular mediation, this pathway exerts positive regulatory effects on M2 macrophage polarization. The downstream Akt/mTORC1 signal can obviously upregulate the expression of M2 characteristic markers such as Arg1 and Mrc1, and maintain the unique metabolic characteristics of M2 macrophages dominated by fatty acid oxidation and mitochondrial respiration, while it shows inhibitory effects on M1 polarization process.
Activated under metabolic stress conditions or mediated by upstream LKB1 kinase, AMPK can inhibit mTORC1 signal activity, maintain mitochondrial biosynthesis and intracellular oxidative metabolism level, and serve as an important metabolic checkpoint to promote macrophages to form M2 polarization phenotype.
The IL-6-JAK-STAT3 signaling pathway is closely related to the maintenance of stable M2 cell phenotype, and targeted inhibition of this pathway can significantly facilitate M1-type polarization tendency of macrophages.
Current research on macrophage phenotypic identification has gone beyond simple detection of limited marker molecules. Combined with multi-omics technology, high-precision imaging means and biophysical detection methods, researchers can achieve more accurate typing identification of polarized macrophages with the technical support of ExKits complete detection solutions.
Macrophages occupy the core position in the whole immune response network, and exert dual regulatory effects in human physiological steady state maintenance and disease occurrence and development. They can not only initiate and amplify inflammatory responses, but also actively promote inflammatory regression and participate in tissue damage repair processes.
Most tumor-associated macrophages (TAMs) present M2-like pro-tumor polarized characteristics, which can mediate local immune suppression, accelerate angiogenesis and promote tumor distant metastasis. Nevertheless, relevant studies have found that some malignant tumors such as triple-negative breast cancer can induce macrophages to form mixed M1/M2 intermediate phenotype, which still shows obvious tumor-promoting effects. In addition, partial M1-like tumor infiltrating macrophages also participate in accelerating the progression of oral squamous cell carcinoma under specific pathological backgrounds. Therefore, the core goal of tumor immunotherapy is to induce the phenotypic transformation of TAMs from M2 type to M1 type so as to rebuild effective anti-tumor immune response.
The pathological process of rheumatoid arthritis is accompanied by dominant M1/Th1-type immune response activation. Massive pro-inflammatory cytokines and matrix metalloproteinases secreted by M1 macrophages directly induce joint tissue destruction. Promoting the shift of immune balance towards M2/Th2 direction can effectively alleviate inflammatory lesions and facilitate disease remission. Similarly, M1 macrophages can aggravate atherosclerotic plaque instability, while M2 macrophages are conducive to inflammatory lesion repair. Multiple active substances represented by ELABELA have been verified to exert therapeutic effects by regulating M1/M2 balance.
The orderly transformation process from M1 inflammatory phenotype to M2 repair phenotype is the key prerequisite for normal wound healing. However, sustained excessive activation of M2 macrophages or abnormal emergence of specific fibrotic macrophage subsets can induce irreversible fibrotic lesions in liver, lung and other organs. On the contrary, in necrotizing enterocolitis related research, exogenous supplementation of M2 macrophage derived extracellular vesicles can effectively induce local macrophage M2 polarization and relieve excessive intestinal inflammatory damage.
In chronic liver injury models, omega-3 unsaturated fatty acids and their derived specialized inflammatory resolution mediators can induce hepatoprotective M2 macrophage polarization, reduce intracellular oxidative stress level and relieve liver local inflammatory infiltration. In retinopathy research, macrophage polarization state shows obvious phase transformation rule: early-stage M1 macrophage mediated inflammatory response promotes neovascularization, while late-stage M2 dominant state is beneficial to abnormal vascular regression, providing clear time window guidance for targeted intervention treatment.
In-depth basic research on macrophage polarization mechanism has laid a solid theoretical foundation for clinical transformation application, and multiple innovative targeted intervention strategies have been gradually developed and verified.
How to accurately deliver regulatory active substances to specific macrophage subgroups in vivo has always been a major difficulty in clinical intervention research. Modified nanocarriers such as mannose modified liposomes and targeted polypeptide modified ferritin nanocages can efficiently deliver functional small interfering RNA and TLR agonist substances to tumor local M2-type TAMs, successfully realizing the phenotypic reprogramming from immune suppressive M2 phenotype to anti-tumor M1 phenotype.
A variety of natural plant-derived active components have been confirmed to possess excellent macrophage polarization regulatory activity. For instance, codonopsis pilosula derived glycopeptide components can reverse M2-type TAMs polarization state through regulating intracellular metabolic level and core signaling pathways. In addition, myricetin can relieve the pathological symptoms of atopic dermatitis by inhibiting excessive M1 inflammatory response and promoting M2-type immune balance restoration via regulating JAK-STAT and NF-κB signaling pathways.
Macrophage derived bioactive vesicles have become a new research hotspot in immune regulation field. Exogenous administration of M2 macrophage derived nanovesicles can effectively promote the transformation of local inflammatory M1 macrophages into immune regulatory phenotype, which has achieved good intervention effect in inflammatory bowel disease models. Meanwhile, blocking tumor cell derived exosomes carrying polarization related signal molecules can cut off the M1-to-M2 polarization induction pathway and inhibit the formation of tumor pre-metastatic microenvironment in distant organs.
Different polarized macrophage subtypes have completely independent intracellular metabolic modes: M1 macrophages mainly rely on aerobic glycolysis to provide energy, while M2 macrophages are dominated by oxidative phosphorylation metabolism. Targeted regulation of core metabolic pathways such as intracellular phosphate ion transport and AMPK-mTOR signal axis has gradually become an important new direction to regulate macrophage polarization tendency.
Macrophages are highly flexible immune effector cells, and their functional differentiation into pro-inflammatory M1 phenotype or repair-type M2 phenotype plays a decisive role in maintaining human body physiological homeostasis and determining disease development trend. Regulated by core signaling pathways including JAK-STAT and NF-κB, the dynamic balance of macrophage polarization state participates in the whole pathological process of tumor occurrence, chronic inflammatory lesion formation and tissue damage repair. Under pathological conditions, this balance is often broken, such as the massive accumulation of immune suppressive M2-like macrophages in tumor tissues leading to local immune escape phenomenon.
At present, mainstream therapeutic research mainly focuses on utilizing targeted nanocarrier delivery system, natural active regulatory factors and metabolic remodeling means to realize directional reprogramming of macrophage polarization phenotype. In the future, relevant research will gradually break through the limitations of traditional simple M1/M2 binary classification theory. Combined with high-precision single-cell sequencing technology, researchers will further clarify the complex heterogeneous characteristics of macrophage populations, and develop precise intervention schemes adapting to different disease microenvironments, so as to dynamically regulate macrophage immune function, restore body immune homeostasis and improve overall clinical treatment efficacy with the full set of technical solutions provided by ExKits.
[1] Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization [J]. Eur J Pharmacol. 2020 Jun 15;877:173090.
[2] Cutolo M, Campitiello R, Gotelli E, Soldano S. The Role of M1/M2 Macrophage Polarization in Rheumatoid Arthritis Synovitis [J]. Front Immunol. 2022 May 19;13:867260.
[3] Zhang X, Li J, Liu Y, et al. Myricetin alleviates DNCB-induced atopic dermatitis by modulating macrophage M1/M2 polarization [J]. Int Immunopharmacol. 2025 Oct 10;163:115212.
[4] Hung CH, Hsu HY, et al. Arsenic Induces M2 Macrophage Polarization and Shifts M1/M2 Cytokine Production via Mitophagy [J]. Int J Mol Sci. 2022 Nov 10;23(22):13879.
[5] Strizova, Z., Benesova, I., et al. (2023). M1/M2 macrophages and their overlaps – myth or reality [J]? Clinical Science (London, England: 1979), 137(15), 1067.
[6] Wang, N., Liang, H., & Zen, K. (2014). Molecular Mechanisms That Influence the Macrophage M1–M2 Polarization Balance [J]. Frontiers in Immunology, 5, 614.
[7] Li J, Yu S, et al. The phase changes of M1/M2 phenotype of microglia/macrophage following oxygen-induced retinopathy in mice [J]. Inflamm Res. 2021 Feb;70(2):183-192.
[8] Pagie, S., Gérard, N., & Charreau, B. (2018). Notch signaling triggered via the ligand DLL4 impedes M2 macrophage differentiation and promotes their apoptosis [J]. Cell Communication and Signaling: CCS, 16, 4.
[9] Miki, S., Suzuki, J. I., et al. (2021). S-1-Propenylcysteine promotes IL-10-induced M2c macrophage polarization through prolonged activation of IL-10R/STAT3 signaling [J]. Scientific Reports, 11, 22469.
[10] Sun X, Li Z, Wang X, He J, Wu Y. Inorganic Phosphate as "Bioenergetic Messenger" Triggers M2-Type Macrophage Polarization [J]. Adv Sci (Weinh). 2024 Apr;11(13):e2306062.
[11] Liu H, Yao M, Ren J. Codonopsis pilosula-derived glycopeptide dCP1 promotes the polarization of tumor-associated macrophage from M2-like to M1 phenotype [J]. Cancer Immunol Immunother. 2024 May 14;73(7):128.
[12] Peng, Y., Zhou, M., et al. (2023). Regulatory Mechanism of M1/M2 Macrophage Polarization in the Development of Autoimmune Diseases [J]. Mediators of Inflammation, 2023, 8821610.
[13] Liu L, Stokes JV, Tan W, Pruett SB. An optimized flow cytometry panel for classifying macrophage polarization [J]. J Immunol Methods. 2022 Dec;511:113378.
[14] Selig, M., Poehlman, L., Lang, N. C., Völker, M., Rolauffs, B., & Hart, M. L. (2024). Prediction of six macrophage phenotypes and their IL-10 content based on single-cell morphology using artificial intelligence [J]. Frontiers in Immunology, 14, 1336393.
[15] Chow, L., Soontararak, S., Wheat, W., Ammons, D., & Dow, S. (2022). Canine polarized macrophages express distinct functional and transcriptomic profiles [J]. Frontiers in Veterinary Science, 9, 988981.
[16] Siniyeh, A. A., Alshaer, W., et al. (2025). Comparative analysis of RT-qPCR, flow cytometry, and Di-4-ANEPPDHQ fluorescence for distinguishing macrophage phenotypes [J]. Biochemistry and Biophysics Reports, 44, 102225.
[17] Xue, J., Schmidt, S. V., et al. (2014). Transcriptome-Based Network Analysis Reveals a Spectrum Model of Human Macrophage Activation [J]. Immunity, 40(2), 274.
[18] Liu, B., Nguyen, P. L., Li, P., Naldrett, M. J., Alvarez, S., & Yu, J. (2025). Identification of novel protein biomarkers of macrophage polarization using comparative proteomic analyses of murine primary macrophages [J]. The Journal of Immunology Author Choice, 214(10), 2730.
[19] Vishnyakova, P., Elchaninov, A., Fatkhudinov, T., & Kolesov, D. (2025). Unravelling approaches to study macrophages: From classical to novel biophysical methodologies [J]. PeerJ, 13, e19039.
[20] Coulis, G., Jaime, D., et al. (2023). Single-cell and spatial transcriptomics identify a macrophage population associated with skeletal muscle fibrosis [J]. BioRxiv, 2023.04.18.537253.
[21] Li S, Fu X, Ning D, et al. Colon cancer exosome-associated HSP90B1 initiates pre-metastatic niche formation in the liver by polarizing M1 macrophage into M2 phenotype [J]. Biol Direct. 2025 Apr 15;20(1):52.
[22] Wang Q, Ma W. Revisiting TAM polarization: beyond M1- and M2-type TAM toward clinical precision in macrophage-targeted therapy [J]. Exp Mol Pathol. 2025 Sep;143:143:104982.
[23] Shan H, Dou W, Zhang Y, Qi M. Targeted ferritin nanoparticle encapsulating CpG oligodeoxynucleotides induces tumor-associated macrophage M2 phenotype polarization into M1 phenotype and inhibits tumor growth [J]. Nanoscale. 2020 Nov 12;12(43):22268-22280.
[24] Pe KCS, Saetung R, et al. Triple-negative breast cancer influences a mixed M1/M2 macrophage phenotype associated with tumor aggressiveness [J]. PLoS One. 2022 Aug 12;17(8):e0273044.
[25] Li CX, Gong ZC, Yu JW. Deliberation concerning the role of M1-type macrophage subset in oral carcinogenesis [J]. J Exp Clin Cancer Res. 2024 Aug 8;43(1):220.
[26] Shou X, Wang Y, Jiang Q, Chen J, Liu Q. miR-126 promotes M1 to M2 macrophage phenotype switching via VEGFA and KLF4 [J]. PeerJ. 2023 Mar 31;11:e15180.
[27] Tang L, Yi X, Tan W, et al. ELABELA Ameliorates Atherosclerosis Through Restoring the M1/M2 Macrophage Balance in ApoE(-/-) Mice [J]. J Am Heart Assoc. 2025 Jul 15;14(14):e041261.
[28] Fabre T, Barron AMS, et al. Identification of a broadly fibrogenic macrophage subset induced by type 3 inflammation [J]. Sci Immunol. 2023 Apr 14;8(82):eadd8945.
[29] Liu, C., Gong, J., et al. (2025). M2-type macrophage nanovesicles regulate the inflammatory response after necrotizing enterocolitis by inducing M1 to M2-like macrophage polarization [J]. Frontiers in Cellular and Infection Microbiology, 15, 1664897.
[30] Videla LA, Valenzuela R, Del Campo A, Zúñiga-Hernández J. Omega-3 Lipid Mediators: Modulation of the M1/M2 Macrophage Phenotype and Its Protective Role in Chronic Liver Diseases [J]. Int J Mol Sci. 2023 Oct 24;24(21):15528.